|
|
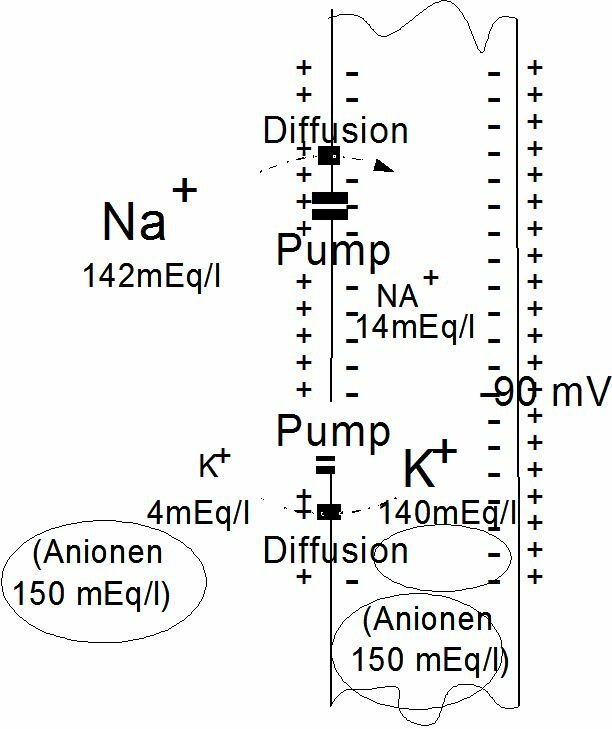
Mechanism of the heart muscleNatalie Diekmann presentationElectrical properties of cell membranesHeart muscle cells contain a contraction-mechanism which is triggered electrically via the so-called action potential. The action potential is an elementary physiological phenomenon that muscle cells and neurons have in common. The action potentials of the heart muscle cells constitute the basis for the electrocardiogram (ECG), which will be discussed later. Note (from Chris) to Jos Spaan: Maybe not relevant for this chapter, but I would like to mention this link http://www.ecglibrary.com/ecghome.html. It could replace figure 4.10 of the syllabus. The composition and particularly the electrolyte composition of the intracellular and extracellular liquid differs substantially. The intracellular liquid contains large amounts of potassium, magnesium and phosphate ions, the extracellular liquid large amounts of sodium and chlorine ions. The cell membrane sees to it that these differences endure, because it makes it as good as impossible for certain substances to permeate. For K+ and Na+ the cell membrane has an active "pumping mechanism", which transports Na+ from the inside to the outside and K+ from the outside to the inside. It is the concentration differences over the membrane of these ions which give cause to the potential difference over the membrane, the membrane potential (see Figure~\ref{Fig:rustpot}). In the stationary case we speak of a rest potential. Every type of cell has its own rest potential, but usually it lies between -75 and -95 mV. The preservation of this rest potential takes energy (oxygen).
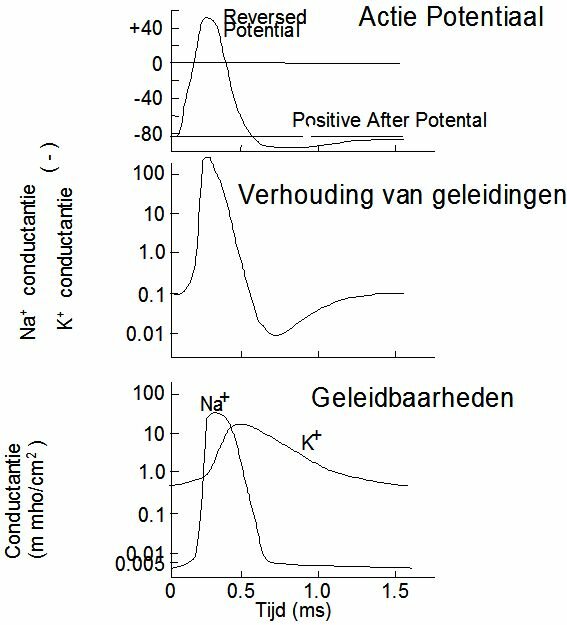
The action potential is this succesion of membrane potential changes, which results after a sudden increase of the membrane's conductibility for sodium. The action potential is finished when the rest potential has established itself again. An action potential arises when the some part of the membrane is stimulated by an electrical, chemical, mechanical or thermal stimulus. In fact almost every disruption of the membrane's state of rest will result in an action potential. The action potential can be divided into two stages: depolarisation and repolarisation. When the conductibility of the pores for Na+ suddenly increases (5000 x), Na+ -ions flow into the cell and can turn the cell's contents positive with respect to the outside (reversal potential). After depolarisation the membrane becomes permeable for Na+ again and the rest potential is restored. The latter process is called repolarisation. At the start of repolarisation the K+ permeability of the membrane increases, as a result of which large amounts of K+ will flow from the cell. Because of this, the cell potential may drop till below the rest potential. The K+ permeability drops again, Na+ is being pumped out of the cell and K+ difuses back in again. The rest potential has been restored. Restoring the rest potential is an active process (Na-K pump) and therefore costs energy, including oxygen.
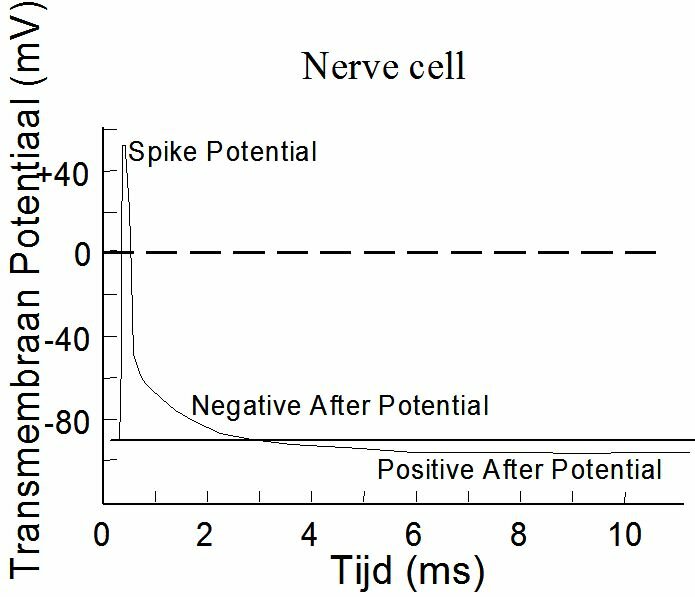
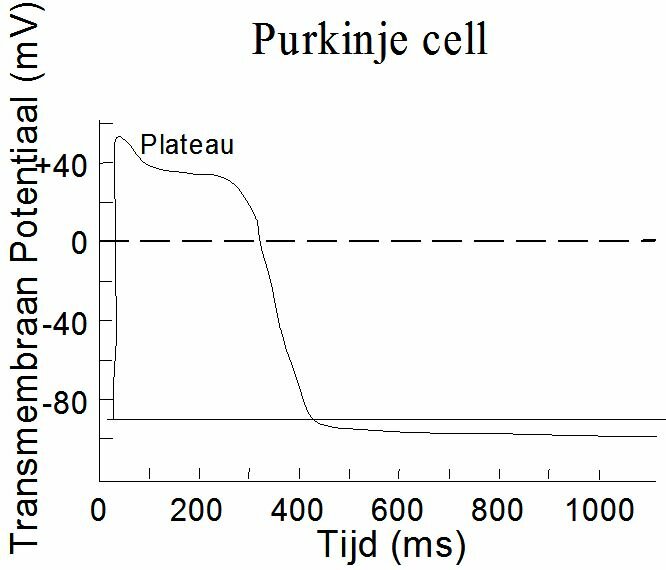
Depolarisation of a membrane starts locally. The ionic concentration gradients cause electrical currents along the membrane which depolarize the adjacent membrane. It is an "all or nothing" principle. The entire membrane depolarizes, or nothing at all. The shape of an action potential depends on the cell. Figure ~\ref{Fig:actiepot} shows the "spike potential" of a neuron and ~\ref{Fig:actiepot2} that of a "Purkinje-fibre" of the heart. The Purkinje-fibres provide the electric pulse conductivity in the heart (Chapter~\ref{ch:elect}). The Purkinje action potential shows a plateau and is much more inert than the spike potential.
The heart muscle cell and its contraction mechanismThe heart muscle cell forms a unit in which chemical energy is converted into mechanical power and/or energy. Before we go and look at the muscle cell in detail, it is good to keep in mind that
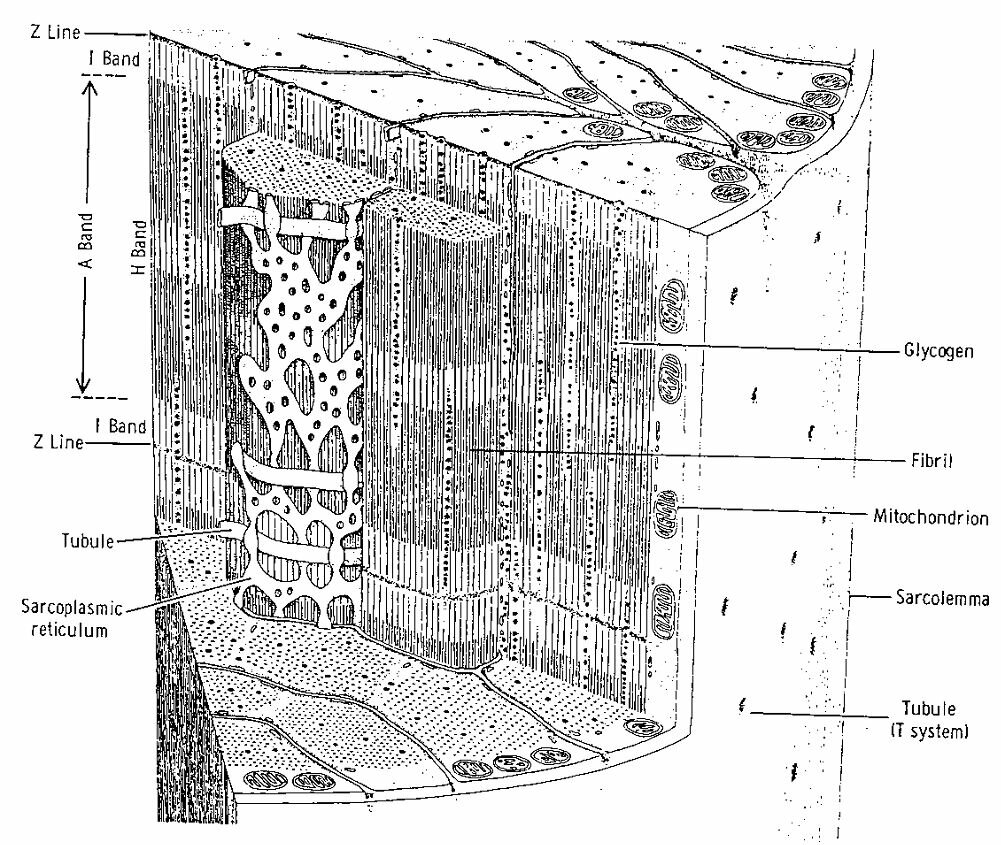
Muscle cells (of the heart muscle and the skeleton muscles, but not of the smooth muscles) are cylindrical, at the tips separated from each other by intercalated discs and cell membranes called sarcolemma (Figure ~\ref{Fig:musclefiber}). The sarcolemma has bulges at regular intervals. These are the t-tubuli. Both sarcolemma and t-tubuli generate action potentials. Every cell contains many mitochondria} where ATP (Adenosine Tri-Phosphate) is made. ATP serves as fuel for the contraction. The fibrils or myofibrils form the cell's contractile elements. They are surrounded by the sarcoplasmic reticulum. This is a complex network of little tubes which lies pushed against the t-tubuli at many places. The sarcoplasmic reticulum is involved in the initiation of the contraction, and serves as storage for calcium.
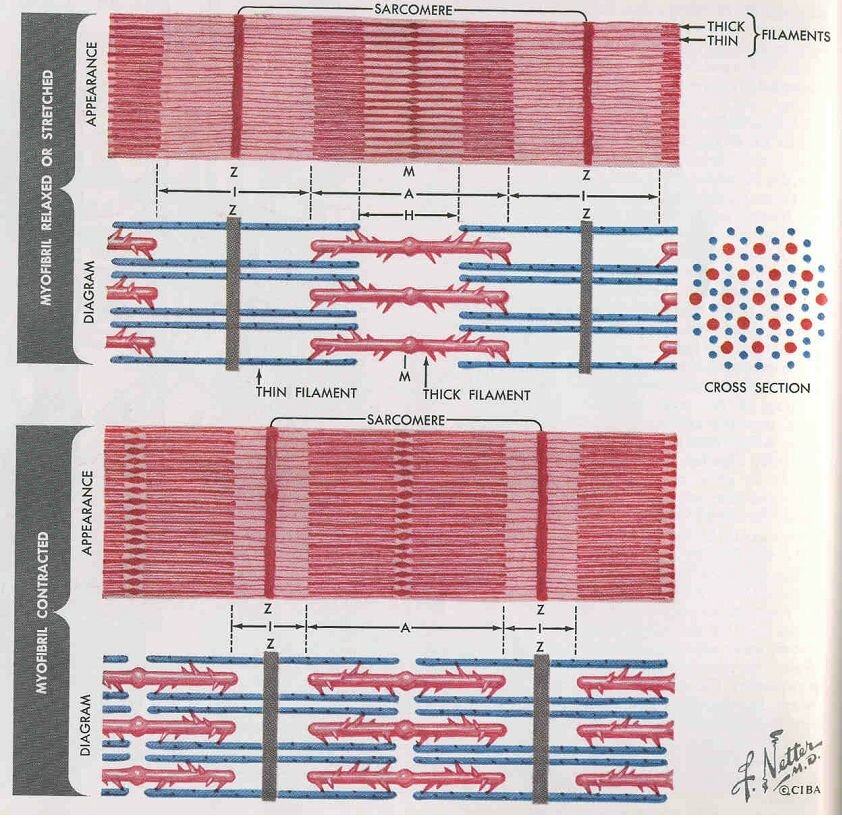
Myofibrils are composed of smaller units called sarcomeres. This well-ordered structure becomes clear from the electron microscopic images (see Figure~\ref{Fig:electron}). The separations between sarcomeres are called Z-lines. Hence, every sarcomere is bounded by 2 Z-lines. The sarcomere itself consists of series of 2 types of structures called {\bf thin and thick filaments}, or {\bf actin- and myosin filaments}. Every thin filament is equidistant from three thick filaments (see Figure ~\ref{Fig:sarcomeer}). The thick filament consists of a large number of myosin molecules (M = 490 000). Every myosin molecule has one block-shaped tail end, which sticks out from the thick filament in the direction of the thin filament. The myosin tail ends cling together and form the M-line at the electron microscopic images. The thin filament consists mainly of actin molecules that have been polymerized in a double helix.
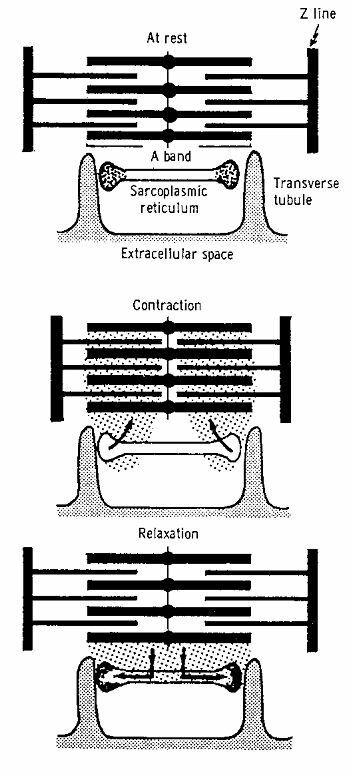
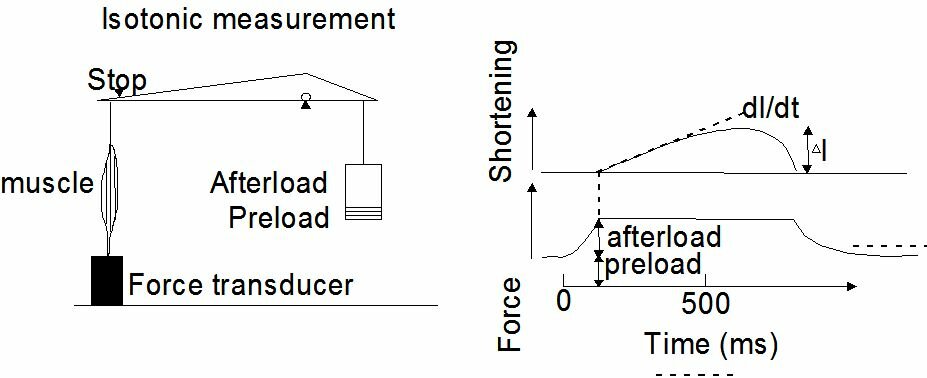
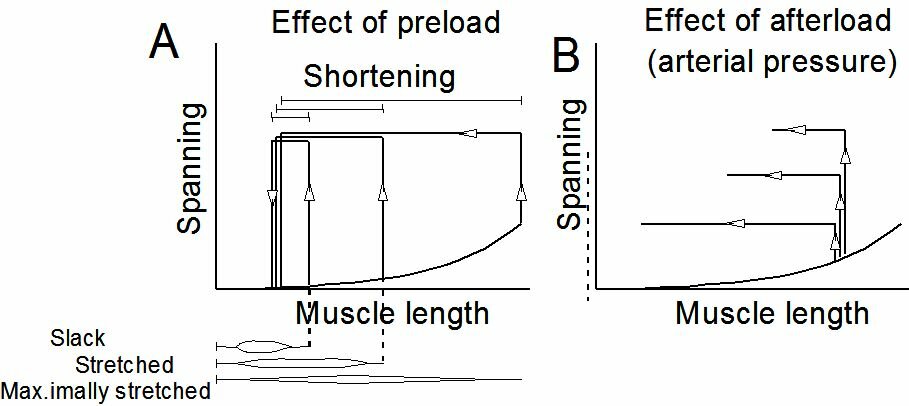
Excitation and contractionThe contraction of the filaments is effectuated by the forming of {\bf cross-bridges} between the myosin heads and specific bonding places of troponin to the actin molecules. The cross-bridges come into being when $Ca^{2+}$ reacts with troponin, which results in the telescoping of the thick and the thin filaments, and thus in a shortening of the fibrils. The contraction process seems to come about as follows (see Figure ~\ref{Fig:musclecontr}). When the heart musle is at rest, there is calcium in the sarcoplasmic reticulum, the sarcolemma, and the extracellular space. Whenever an action potential occurs, this is called excitation of the cell. Due to this excitation, calcium is released from the sarcoplasmic reticulum and sarcolemma. This calcium reacts with troponin (troponin C, to be exact), which results into the coming about of cross-bridges. With the decline of the action potential the $Ca^{2+}$ returns to its storage. The heart muscle relaxes and returns to its state of rest. For the undoing of the cross-bridges, ATP is needed. When the muscle actually shortens itself, it means that for just one contraction several cross-bridges have to be formed (requiring $Ca^{2+}$) and have to be undone (requiring ATP). Mechanics of heart muscle tissueNon-activated heart tissue shows elastic behaviour, albeit not ideally elas\-tic (see Figure~\ref{Fig:preafterload}). Activation of the heart muscle cells causes an internal build-up of tension which, external conditions permitting, will lead to a shortening of the muscle in the longitudinal direction of the fibrils. Contrary to skeleton muscle tissue, heart muscle tissue cannot bring about a tetanus (lasting contraction). A stimulus leads to excitation resulting in a contraction, followed by relaxation. The classic test with which to study the mechanical phenomena of heart muscle tissue, is reflected in Figure~\ref{Fig:contractieproef} a,b. Usually, a papillary muscle or trabecula is used for this test. The strings tightening the mitral valve (valve between a ventricle and an atrium) are connected at their opposite ends to a papillary muscle, which can be pictured as an intraventricular bulge of the heart muscle. Trabeculae are threads of muscle tissue against the inside of the heart muscle. In its non-activated state this little muscle is forced to extend, so that it is prestressed according to the passive force- length relationship. This prestressing is called the "preload". The little muscle can now shorten itself, but first it has to develop the power to do so. (see Figure~\ref{Fig:contractieproef} b). This force is known as the afterload. The results of this test are given in Figure~\ref{Fig:contractieproef}b in terms of tension progress and length change in time. During the time that the muscle length is constant, we speak of isometric contraction, in the time that the muscle power is constant, we speak of isotonic contraction. The defining parameters in the experiment described above are preload and afterload. The variables are time, tension and length. As characterizing parameters of the contraction are used the maximum shortening and the shortening velocity at the moment the muscle tension equals the afterload. When the results are considered in a none too critical way, it turns out that
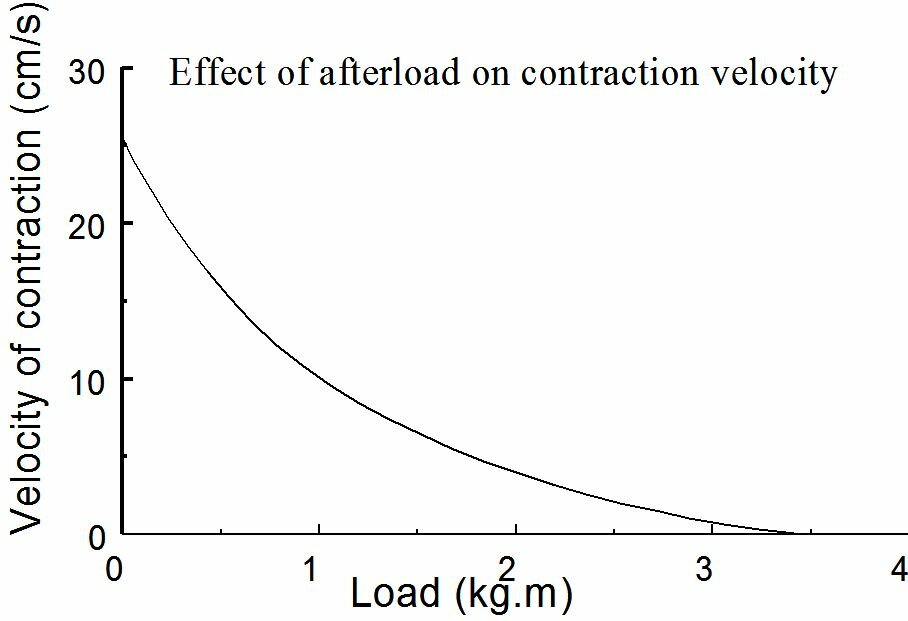
In Figure~\ref{Fig:preafterload} the influence of preload and afterload on the contraction is given in length-tension diagrams. The fact that the shortening velocity decreases when the afterload increases is in accordance with everyone's experience that an object can be lifted less quickly the heavier it is.
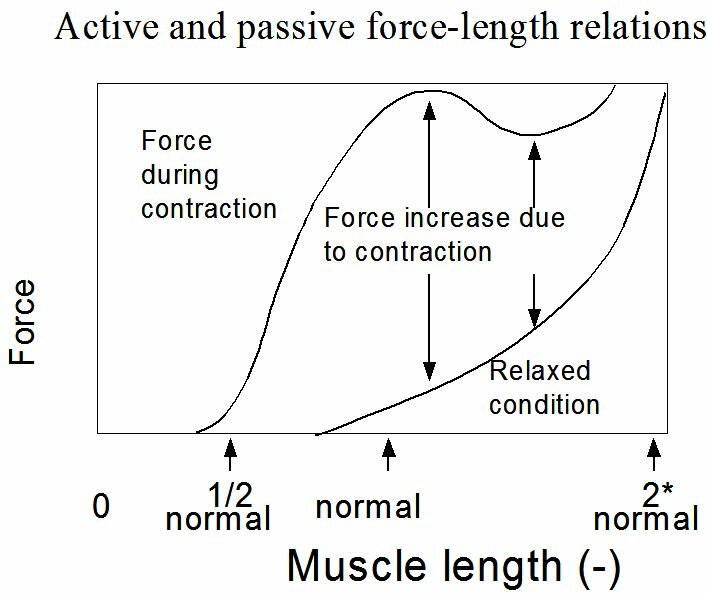
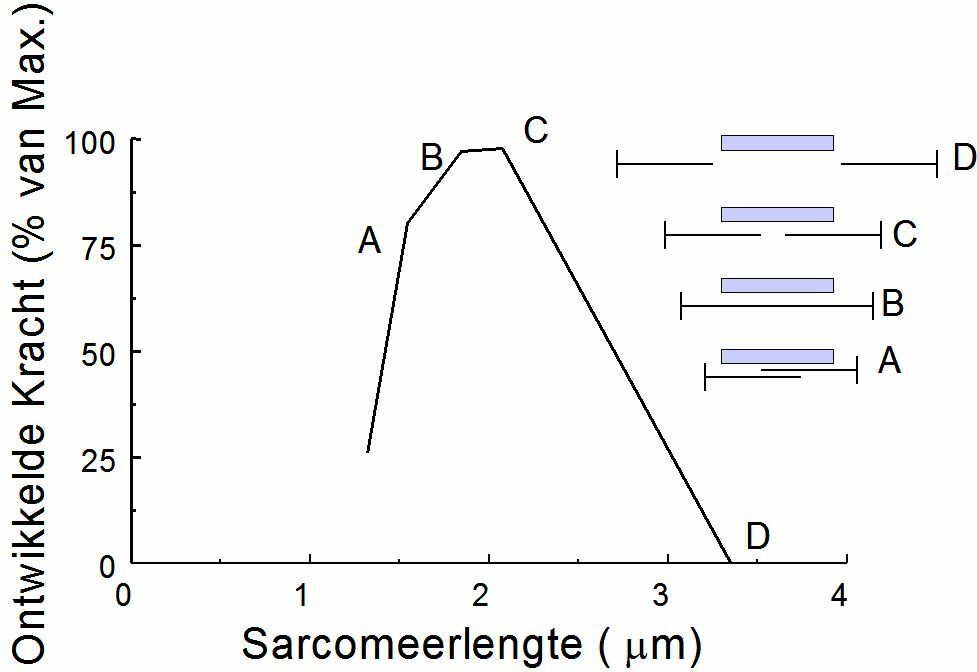
The contraction velocity also depends on the afterload. Without afterload, the contraction velocity reaches a maximum. In case of isometric contraction the contraction velocity equals zero (Figure~\ref{Fig:contractiesnelheid}). When we put the passive and active force-length relationship of a striated muscle into a chart this results in the situation as given in Figure~\ref{Fig:actieve}. By the active force we mean the peak force during an isometric contraction. The active force is the difference between the force on the muscle in activated and passive state at the same sarcomere length. The shape of the active force-sarcomere length curve is given in Figure ~\ref{Fig:krachtlengte}. An optimum is reached at a sarcomere length of approximately 2.2 $\mu$m. At lesser sarcomere lengths the force decreases because the filaments are pressed into each other. At larger lengths the force decreases because the filamentary overlap decreases. Actually, when the heart muscle is stretched to the extent that it is causing the active force to drop, the muscle cells will get heavily damaged. This stands in great contrast with the striated muscles.
A very important defining parameter has not been mentioned yet, viz. the muscle's intrinsic contraction properties. (see also http://www.biophysics.org/btol/muscle.html) From the description of the contraction mechanism it is obvious that without $Ca^{2+}$ there is not going to be a contraction. These intrinsic properties of the heart muscle are referred to as contractility. This contractility can be influenced by various pharmaceuticals. A contractility increase will lead to larger shortenings at a constant preload and afterload. We will come back to the concept of contractility when discussing the heart function. (Chapter~\ref{ch:pomp}).
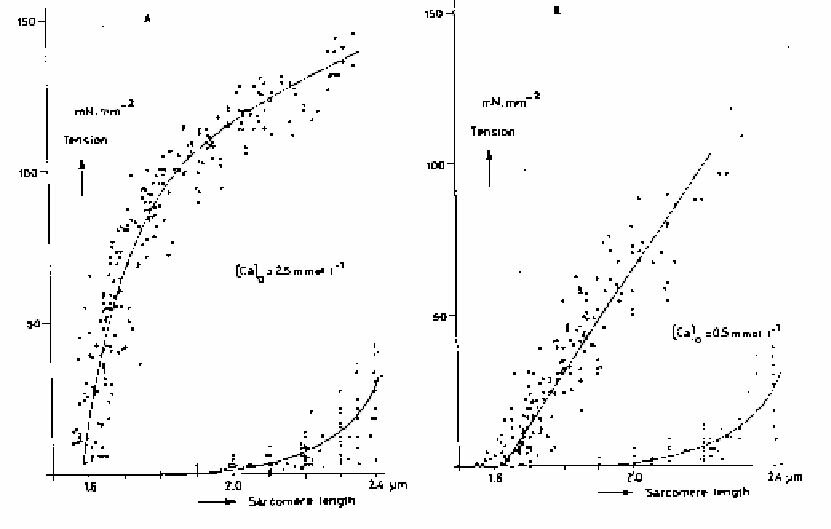
When reading literature on this subject, the reader should be critical as to the experimental conditions applied. When a muscle is clamped, the part between and close to the clamps will be destroyed. The sarcomeres will no longer shorten themselves in this part of the muscle. This destroyed part of the muscle does still retain its elasticity, however. Contraction of the healthy part of the muscle yields a lengthening of the destroyed part. When the total test strip length remains constant, sarcomere shortening will indeed occur in the centre. Sarcomere length can be measured continuously by sending a laser beam through the muscle. The ordened structure of the sarcomeres provides a grid and an interference pattern is obtained from which the bar distance of the grid (sarcomere length) can be calculated. Via a feedback system an arbitrary shortening can be forced on the muscle. Figure~\ref{Fig:leiden} shows test results obtained with this line-up. The maximum force development during isometric contractions has been plotted against the sarcomere length. Figures~\ref{Fig:leiden}a and b show the passive force-length relationships. In Fig. a the contractility of the muscle is higher than in Fig. b because of what is called a higher activation of the muscle by a higher extra-cellular Ca2+ concentration.
Summary of links: The fibrils or myofibrils form the cell's contractile elements http://www.biophysics.org/btol/muscle.html
|